A few years ago, psychologist Sherman Lee experienced a “grief attack”. When he set out to study them, he discovered just how powerful and all-consuming grief can be


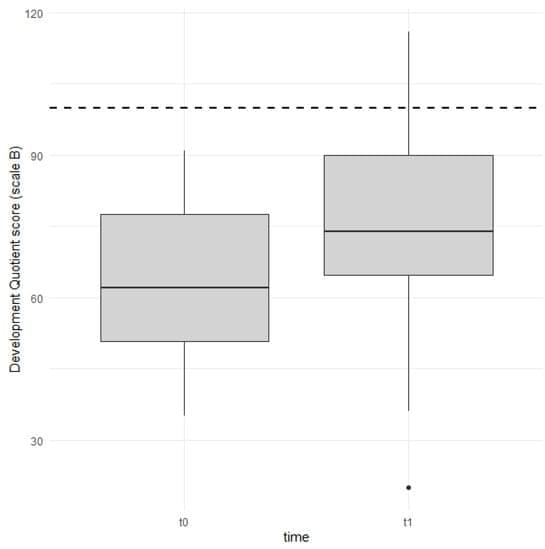
Background/Objectives: The primary objective of the present study was to investigate early global development in children after one year of cochlear implant (CI) use. The secondary objective was to investigate the role of variables such as age at CI activation, gender, and parental schooling in early global development in children with a CI. Methods: The study sample included 24 subjects. All children were affected by severe-to-profound congenital bilateral sensorineural hearing loss (HL). The HL was diagnosed between 1 and 23 months of age (median 3 months) and participants underwent cochlear implant activation at 9–25 months (median 14 months). Participants were evaluated before CI surgery and after one year of CI use using the Italian version of the Griffiths III scales.



Over the past two decades, a powerful but highly specialized branch of control engineering—known as negative imaginary (NI) systems theory—has quietly evolved into a key tool for stabilizing complex, vibration-prone systems, from flexible structures to advanced robotics.
Now, a new study reveals how fast and in what direction this field is growing. By analyzing more than 400 scientific publications from 2004 to 2024 across the world’s leading academic databases, the researchers uncovered a clear trend: NI systems are no longer just a theoretical concept. The work is published in the International Journal of Systems Science.
The field is expanding rapidly, with increasing global participation and a strong shift toward real-world applications such as multi-robot coordination, aerospace systems and nanotechnology.

Large language models (LLMs), the computational models that underpin conversational agents such as Gemini and ChatGPT, are now widely used by people worldwide to rapidly find information, summarize documents and generate texts for specific purposes. Some computer scientists are now combining two or more of these models to create multi-agent systems, which prompt multiple artificial intelligence (AI) agents to interact, cooperate and/or compete with the goal of completing specific tasks.
In some scenarios, however, AI agents could have different objectives and might have access to more or less information than the other agents they are interacting with. Understanding how AI agents typically behave in these situations could help shed more light on the potential benefits and risks of multi-agent systems.
Researchers at Mila, Université de Montréal and McGill University recently set out to explore how the hidden goals of individual AI agents could influence a multi-agent system’s performance, using a framework inspired by the multiplayer social deduction game Werewolf.

One of the most iconic visual representations in biological science is the double helix – the twisting, twin-stranded ladder that defines the shape of DNA molecules.
But not all DNA follows this same recognizable pattern. Alternative kinds of DNA structures do exist, and new technologies are helping to reveal them.
In a study last year, scientists identified where these “non-canonical” forms of DNA (aka non–B DNA) emerge in human and other primate genomes.
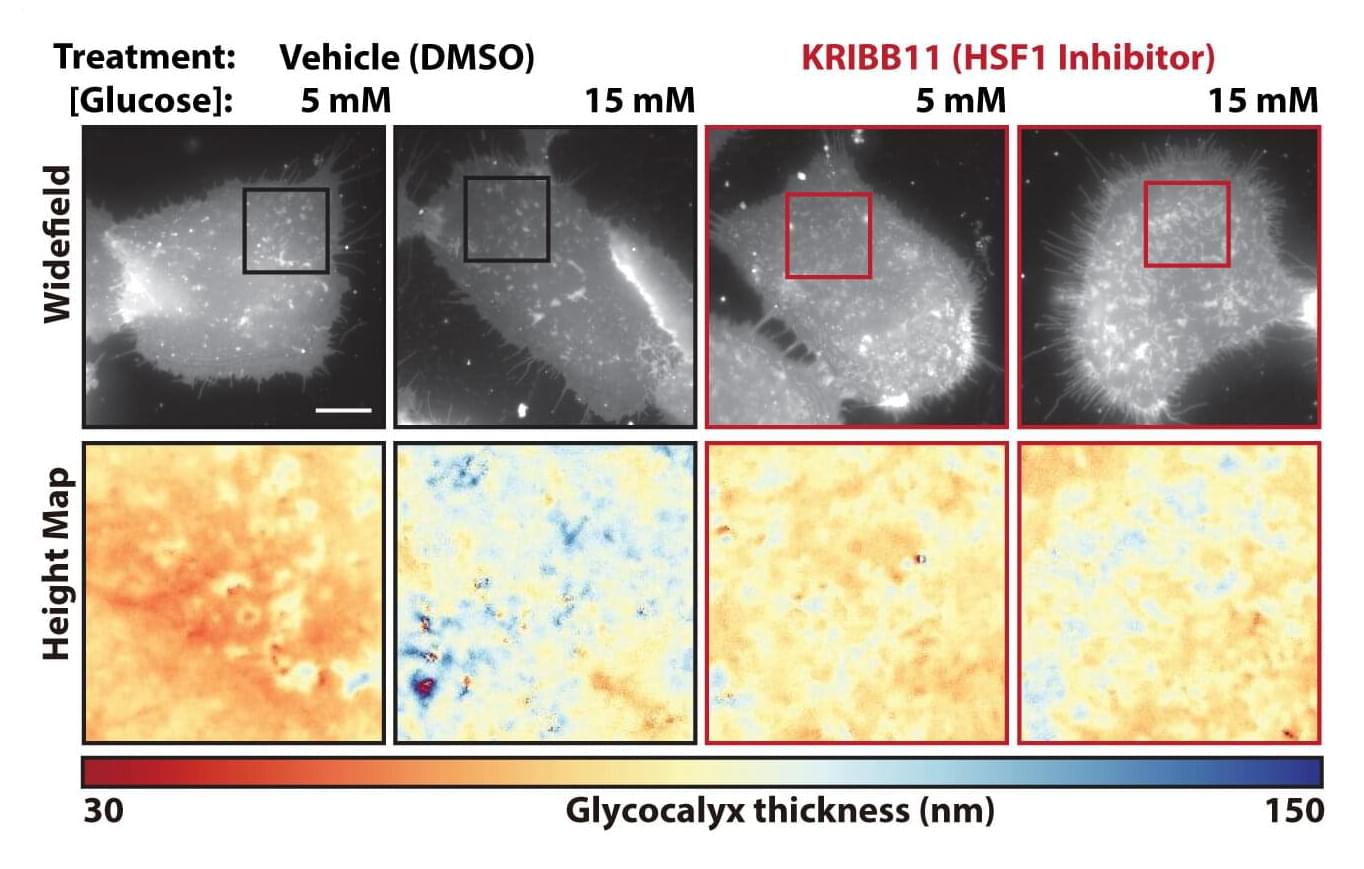
Like spies evading detection by mastering disguises, many cancer cells are adorned with a copious coat of sugar-derived molecules that throws the proverbial hounds of the immune system off the scent.
Scientists at Sanford Burnham Prebys Medical Discovery Institute and collaborators across North America published findings Aug. 7, 2026, in Science Advances showing that cancer cells’ cloaking costumes can result from changes in the nearby neighborhood of immune cells, connective tissue, blood vessels, proteins and carbohydrates called the tumor microenvironment. The researchers also found a way to thin this sugary shroud, enabling cancer cells to be recognized and eliminated by the immune system.
Lead and corresponding author Kevin Tharp, Ph.D., knew that cells squeezed by their surroundings change their mitochondrial function in surprising ways. He realized that a key place where cells would experience this kind of physical pressure was in the tumor microenvironment.