#theoryofeverything #consciousness #philosophy Hey Every0ne, happy new year! Tonight we will discuss a talk on Theories of Everything @TheoriesofE…
Get the latest international news and world events from around the world.

Intelligence on Earth Evolved Independently at Least Twice
Humans tend to put our own intelligence on a pedestal. Our brains can do math, employ logic, explore abstractions, and think critically. But we can’t claim a monopoly on thought. Among a variety of nonhuman species known to display intelligent behavior, birds have been shown time and again to have advanced cognitive abilities. Ravens plan for the future, crows count and use tools, cockatoos open and pillage booby-trapped garbage cans, and chickadees keep track of tens of thousands of seeds cached across a landscape. Notably, birds achieve such feats with brains that look completely different from ours: They’re smaller and lack the highly organized structures that scientists associate with mammalian intelligence.
“A bird with a 10-gram brain is doing pretty much the same as a chimp with a 400-gram brain,” said Onur Güntürkün, who studies brain structures at Ruhr University Bochum in Germany. “How is it possible?”
Researchers have long debated about the relationship between avian and mammalian intelligences. One possibility is that intelligence in vertebrates—animals with backbones, including mammals and birds—evolved once. In that case, both groups would have inherited the complex neural pathways that support cognition from a common ancestor: a lizardlike creature that lived 320 million years ago, when Earth’s continents were squished into one landmass. The other possibility is that the kinds of neural circuits that support vertebrate intelligence evolved independently in birds and mammals.
Dave Pascoe Presents His Blood Test Results (41 Biomarkers)
Join us on Patreon! https://www.patreon.com/MichaelLustgartenPhDDiscount Links/Affiliates: Blood testing (where I get the majority of my labs): https://www.u…

Astronomers Find Far-flung “Super Earths” Are Not Farfetched
A new study shows that planets bigger than Earth and smaller than Neptune are common outside the Solar System.
An international team including astronomers from the Center for Astrophysics | Harvard & Smithsonian (CfA) has announced the discovery of a planet about twice the size of Earth orbiting its star farther out than Saturn is to the sun.
These results are another example of how planetary systems can be different from our solar system.
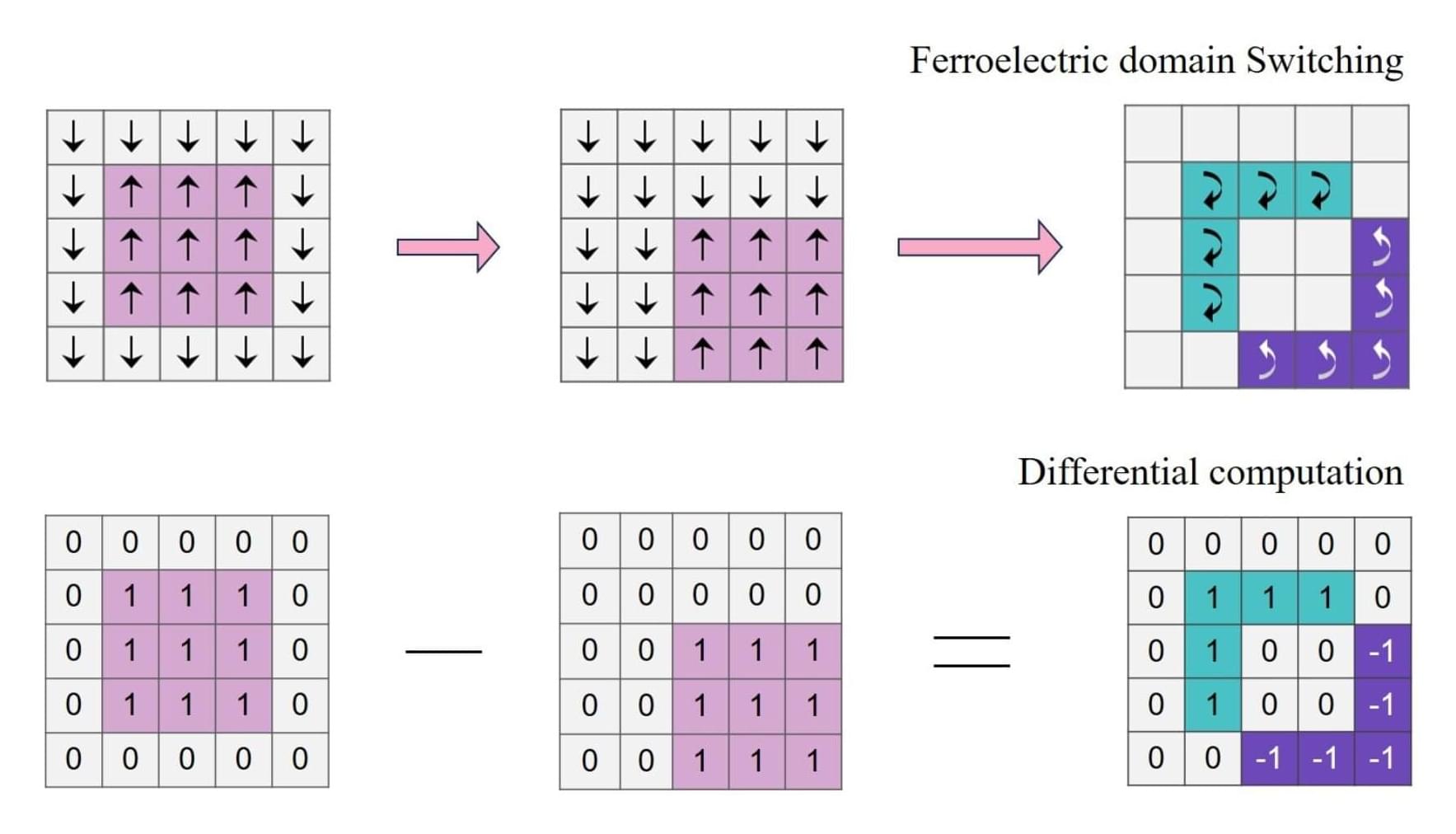
Ferroelectric RAM performs calculations within memory
In a new Nature Communications study, researchers have developed an in-memory ferroelectric differentiator capable of performing calculations directly in the memory without requiring a separate processor.
The proposed differentiator promises energy efficiency, especially for edge devices like smartphones, autonomous vehicles, and security cameras.
Traditional approaches to tasks like image processing and motion detection involve multi-step energy-intensive processes. This begins with recording data, which is transmitted to a memory unit, which further transmits the data to a microcontroller unit to perform differential operations.
Crack-Up Capitalism
A Fortune best nonfiction book of 2023In a revelatory dispatch from the frontier of capitalist extremism, an acclaimed historian of ideas shows how free mark…

Physicists Discover First Room-Temperature 2D Altermagnet
The study identifies a new class of layered antiferromagnets with spin-valley locking, offering efficient spin control without relying on spin–orbit coupling.
Altermagnets are a newly recognized class of materials that show momentum-dependent spin splitting without requiring spin-orbit coupling (SOC) or net magnetization. These materials have recently garnered international attention.
A research team led by Prof. Junwei Liu from the Department of Physics at the Hong Kong University of Science and Technology (HKUST), together with experimental collaborators, published groundbreaking findings in Nature Physics.


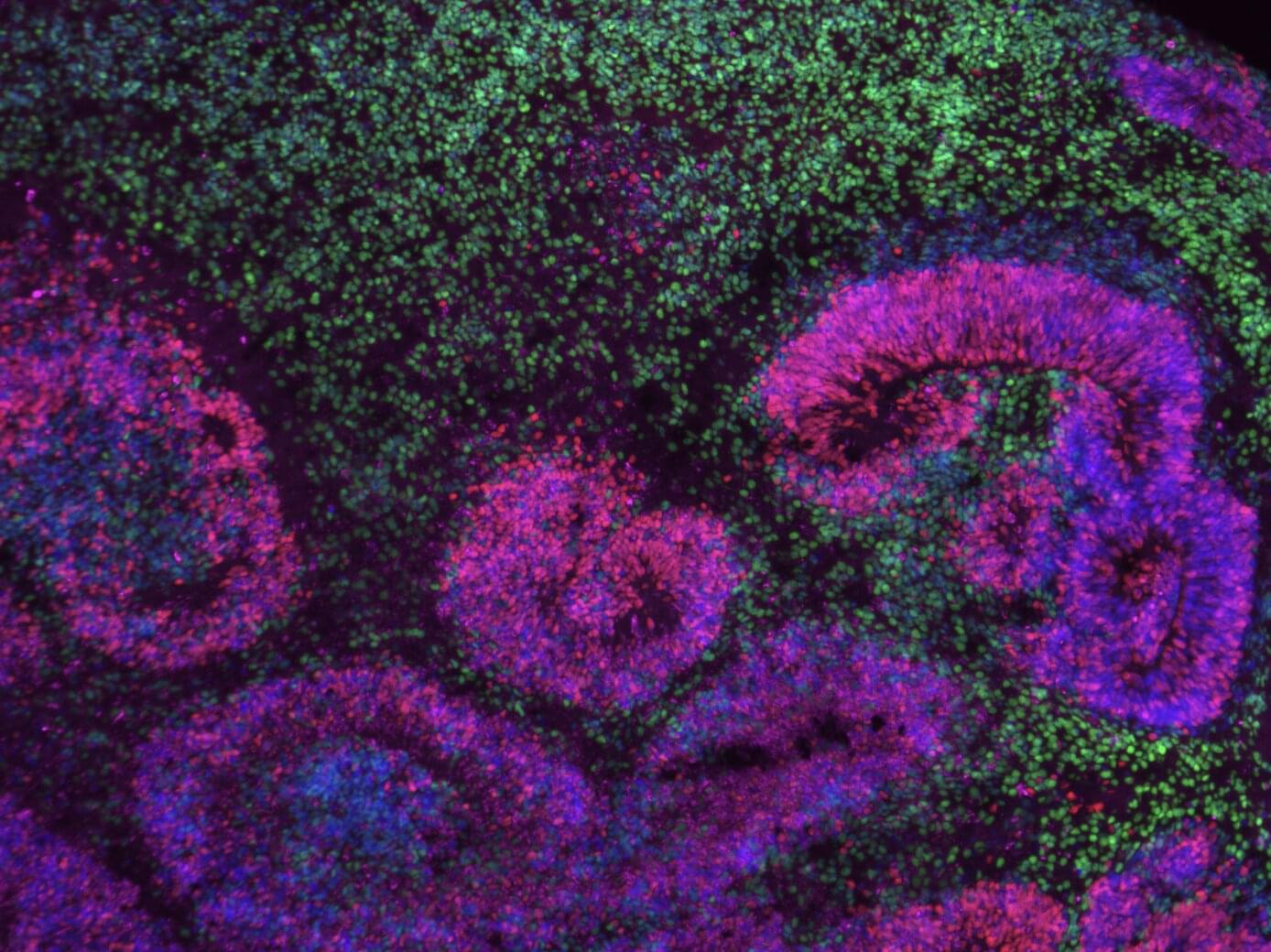
Study uncovers gene networks driving the development of distinct neuron subtypes in the human cerebral cortex
The human brain is known to contain a wide range of cell types, which have different roles and functions. The processes via which cells in the brain, particularly its outermost layer (i.e., the cerebral cortex), gradually become specialized and take on specific roles have been the focus of many past neuroscience studies.
Researchers at the University of California Los Angeles (UCLA) analyzed different datasets collected using single-cell transcriptomics, a technique to study gene expression in individual cells, to map the emergence of different cell types during the brain’s development.
Their findings, published in Nature Neuroscience, unveil gene “programs” that drive the specialization of cells in the human cerebral cortex.