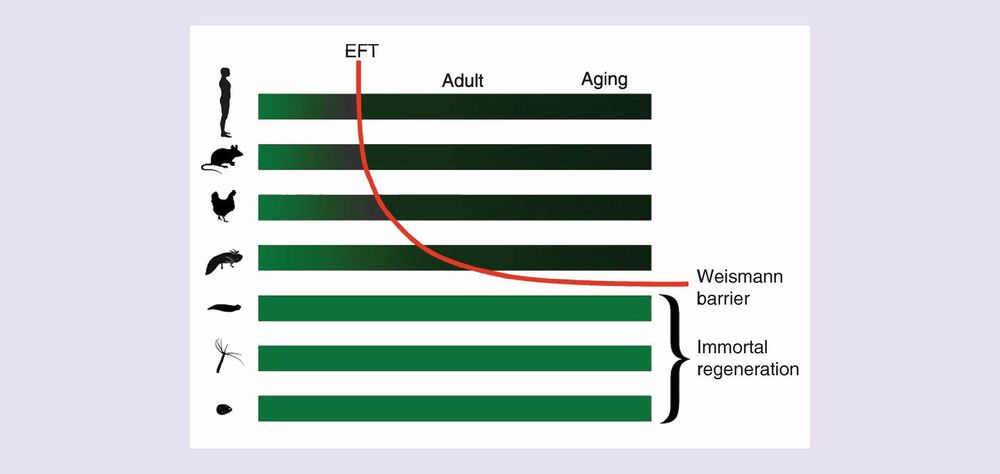
We can evolve bacteria, plants and animals. Is it ethical to evolve the human body? I say yes.
And it becomes a moral imperative because it’s going to be really hard to live on Mars if we don’t fundamentally modify the human body. Right? You go from one cell, mom and dad coming together to make one cell, in a cascade to 10 trillion cells. We don’t know, if you change the gravity substantially, if the same thing will happen to create your body. We do know that if you expose our bodies as they currently are to a lot of radiation, we will die. So as you’re thinking of that, you have to really redesign things just to get to Mars. Forget about the moons of Neptune or Jupiter.
And to borrow from Nikolai Kardashev, let’s think about life in a series of scales. So Life One civilization is a civilization that begins to alter his or her looks. And we’ve been doing that for thousands of years. You’ve got tummy tucks and you’ve got this and you’ve got that. You alter your looks, and I’m told that not all of those alterations take place for medical reasons.
Seems odd.
A Life Two civilization is a different civilization. A Life Two civilization alters fundamental aspects of the body. So you put human growth hormone in, the person grows taller, or you put x in and the person gets fatter or loses metabolism or does a whole series of things, but you’re altering the functions in a fundamental way.