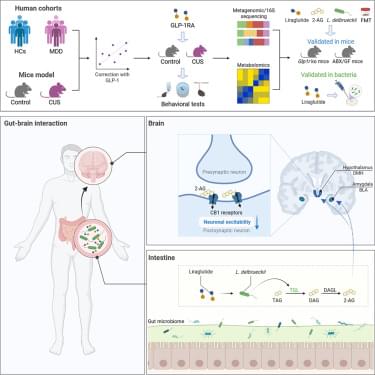
To determine whether canonical GLP-1R signaling is required for liraglutide to remodel the gut microbiota, we performed 16S rRNA sequencing on fecal samples from CUS-exposed wild-type (WT) and Glp1r−/− mice treated with or without liraglutide. Analyses of alpha-diversity, beta-diversity, and genus-level composition revealed that liraglutide changed the microbial structure in CUS mice, although specific compositional shifts differed between WT and Glp1r−/− mice (Figure S6). However, linear discriminant analysis (LDA) identified the genus Lactobacillus as the most significantly enriched taxon following liraglutide treatment in both WT and Glp1r−/− mice (Figures 2 H and 2I). Consistent with this finding, the abundance of Lactobacillus, which was reduced by CUS, was restored by liraglutide in both WT and Glp1r−/− mice (Figure 2 J). To identify the specific Lactobacillus species affected, we performed metagenomic sequencing on fecal samples from CUS mice treated with liraglutide. The Venn diagram showed that L. delbrueckii emerged as the most markedly altered species following liraglutide intervention in CUS mice (Figures 2 K and 2L). Targeted qPCR further validated that CUS-induced reduction in L. delbrueckii abundance was restored by liraglutide treatment in both WT and Glp1r−/− mice (Figures S7 A and S7B). Moreover, semaglutide, another GLP-1R agonist, similarly reversed the CUS-induced reduction of L. delbrueckii, suggesting a shared effect within this class of drugs (Figure S7 C). Together, these results demonstrated that liraglutide enriches intestinal L. delbrueckii in a manner that does not require canonical GLP-1R signaling. Notably, subcutaneous administration of liraglutide reached the gut lumen, and L. delbrueckii was most abundant in the ileum (Figure S8), supporting the in vivo relevance of the proposed mechanism.
To establish the causal role of liraglutide-induced microbial remodeling in mediating its behavioral effects, we performed fecal microbiota transplantation (FMT) from either untreated CUS or liraglutide-treated CUS donors into ABX-pretreated CUS recipients (Figure 2M). Recipients colonized with microbiota from liraglutide-treated donors exhibited significant improvements in depressive-like behaviors, as evidenced by increased sucrose preference in the SPT and reduced immobility in both the TST and FST, whereas microbiota from untreated CUS donors produced no significant behavioral change (Figures 2N–2P). Additionally, we found that FMT from liraglutide-treated donors similarly ameliorated depressive-like behaviors in lipopolysaccharide (LPS)-exposed recipients (Figure S9). We further quantified L. delbrueckii abundance in recipient feces and found that FMT from liraglutide-treated donors elevated L. delbrueckii abundance in recipients (Figure 2Q). Notably, the abundance of L.