{kind=link}
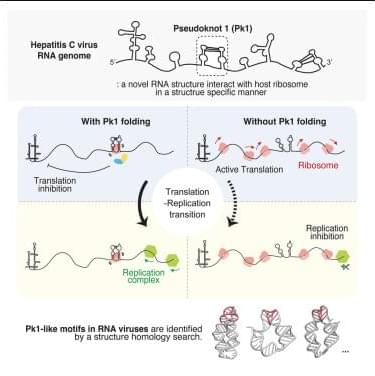
Given that pk1 is located within the HCV coding region, we next investigated a potential role in translation elongation. We inserted pk1 into the ORF of Gaussia luciferase and used the HCV IRES to initiate translation (Figure 3 B, left). To ensure that the highly structured HCV IRES did not affect pk1 folding, we included a 99-nt linker sequence with low structural propensity between the HCV IRES and pk1. As controls, we used the pk1-unzip mutant and a randomized pk1, where the pk1 sequence was shuffled by three nucleotides, thereby maintaining the same amino acid composition as the WT pk1.
The RNA constructs described above were transfected into cells, and luciferase activity was measured at various time points to assess translation activity (Figure 3B, right). We observed that pk1 significantly inhibited translation elongation at all time points tested compared to the pk1-unzip and random controls. Notably, at 4 h post-transfection, there was nearly a 4-fold difference between the activity of pk1 and the pk1-unzip mutant, indicating that the pseudoknot structure itself, rather than just its constituent stems, is crucial for the observed translation inhibition (Figure 3B). To further examine the structural basis of this effect, we introduced a pk1-compensatory mutant that restores the disrupted base pairing in the pk1-unzip mutant using synonymous G-U wobble and A-U base pairs (sequence shown in Figure 3A).