SpaceX conducted 15 of the liftoffs.

For the first time, an international team of scientists has experimentally simulated spontaneous symmetry breaking (SSB) at zero temperature using a superconducting quantum processor. This achievement, which was accomplished with over 80% fidelity, represents a milestone for quantum computing and condensed matter physics.
The results are published in the journal Nature Communications.
The system began in a classical antiferromagnetic state, in which particles have spins that alternate between one direction and the opposite direction. It then evolved into a ferromagnetic quantum state, in which all particles have spins that point in the same direction and establish quantum correlations.
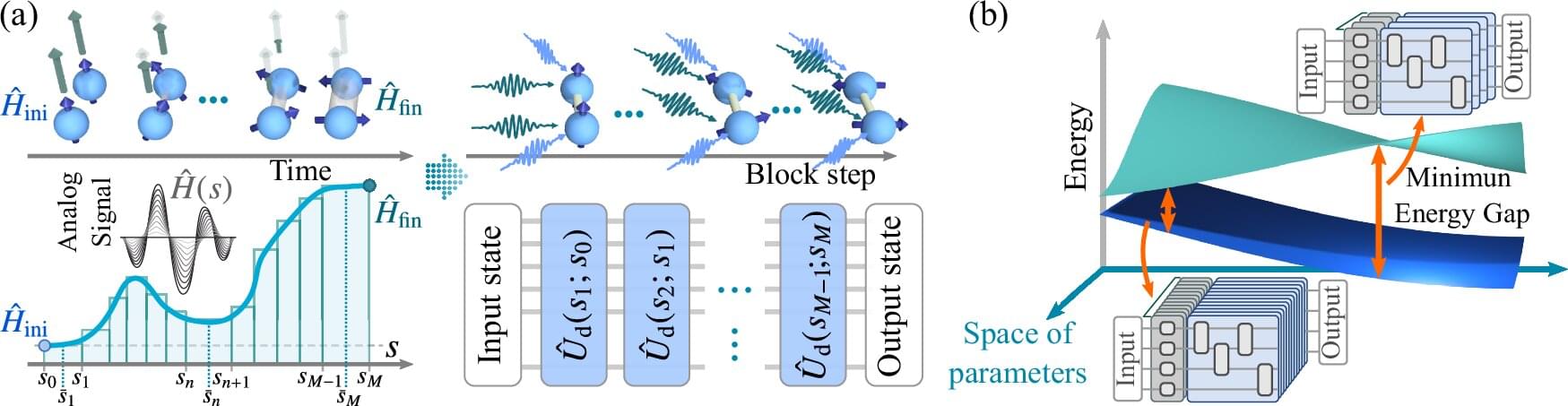
With the rise of 3D printing and other advanced manufacturing methods, engineers can now build structures that were once impossible to fabricate. An emerging design strategy that takes full advantage of these new capabilities is topology optimization—a computer-driven technique that determines the most effective way to distribute material, leading to an optimized design.
Now, a research team including mathematicians from Brown University has developed a new approach that dramatically improves the speed and stability of topology optimization algorithms. The team, a collaboration between researchers at Brown, Lawrence Livermore National Laboratory and Simula Research Laboratory in Norway, detailed their work in two recently published papers in the SIAM Journal on Optimization and Structural and Multidisciplinary Optimization.
“Our method beats some existing methods by four or five times in terms of efficiency,” said Brendan Keith, an assistant professor of applied mathematics at Brown. “That’s a huge computational savings that could enable people to make designs more quickly and inexpensively, or to develop more complex designs with higher resolution.”
Why do some changes in nature unfold gradually, while others occur in the blink of an eye? Rust forming on metal is a slow, steady process that takes days or even weeks to become visible. By contrast, a power grid can collapse in mere seconds. What accounts for this difference?
A research team at Bar-Ilan University has uncovered a surprising mechanism behind these abrupt transitions, a hidden spontaneous sequence of micro-scale events that gradually destabilize a system until it snaps. Their discovery sheds new light on how complex systems behave near critical tipping points and offers a new way to anticipate and perhaps even prevent catastrophic failure.
In their study just published in Nature Communications, the team led by Professors Shlomo Havlin and Aviad Frydman—alongside BIU researchers Ira Volotsenko, Yuval Sallem, and Nahala Yadid, and postdoctoral collaborators Bnaya Gross (Northeastern University) and Ivan Bonamassa (CEU Vienna)—investigated a novel engineered experimental system: interdependent superconducting networks.
Questions to inspire discussion.
🏭 Q: How much LFP cell production capacity does Tesla have in Nevada? A: Tesla’s Nevada facility has equipment for 7–8 GWh of LFP cell production across two production lines, potentially for EVe and grid storage cells.
Tesla Business and Sales.
📊 Q: What are the expectations for Tesla’s Q2 PND report? A: Troy Teslike estimates 356,000 deliveries, while analyst consensus is 385,000, but PND reports are becoming less significant for Tesla’s business model.
💰 Q: What’s crucial for Tesla to become a multi-trillion dollar company? A: Unsupervised FSD rollout and Optimus sales at scale are key, not just increased car or megapack sales.
🇨🇳 Q: How are Tesla’s China sales performing? A: Latest week sales were 20,684 units, down 4.9% QoQ and 11% YoY, but year-to-date figures show Tesla China is closing the gap, down only 4.6% YoY.

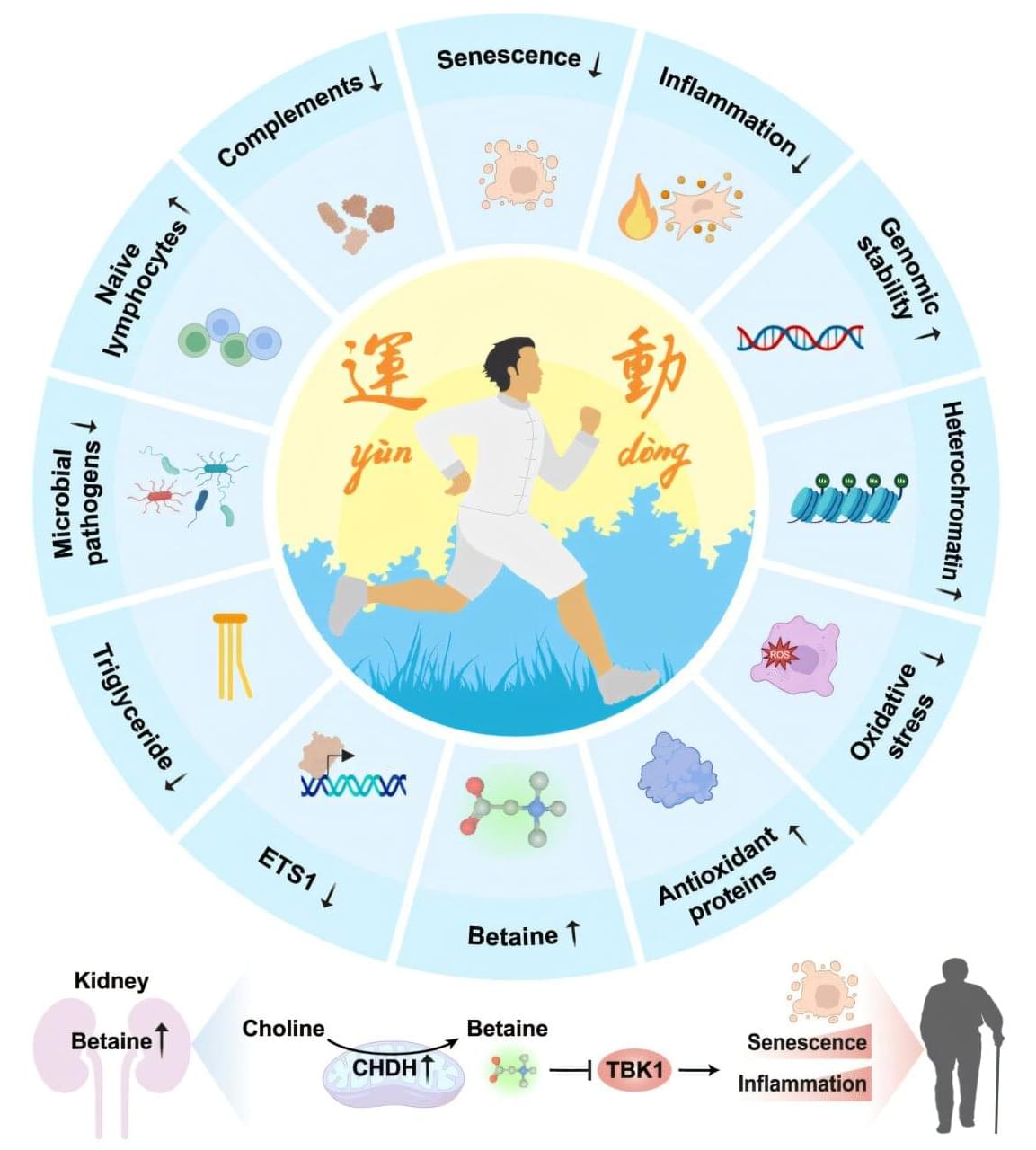
Capital Medical University, in collaboration with the Chinese Academy of Sciences, reports that betaine, a molecule produced in the kidney and enhanced through sustained exercise, operates as a potent inhibitor of inflammatory and aging-related pathways.
Regular physical activity boosts health across cardiovascular, metabolic, and neurological systems. Scientists have traced improvements in immune function, insulin sensitivity, clearing of senescent cells and tissue regeneration to consistent physical activity. Earlier animal studies suggested that long-term exercise can delay aging processes and reduce vulnerability to chronic disease.
Precise molecular explanations for how sustained exercise reshapes human biology remain incomplete. Many investigations have focused on single biomarkers or isolated tissues, leaving a need for systematic maps that can connect exercise to measurable physiological benefits. Specific factors capable of mimicking exercise’s protective effects without requiring continuous physical exertion have remained unclear.
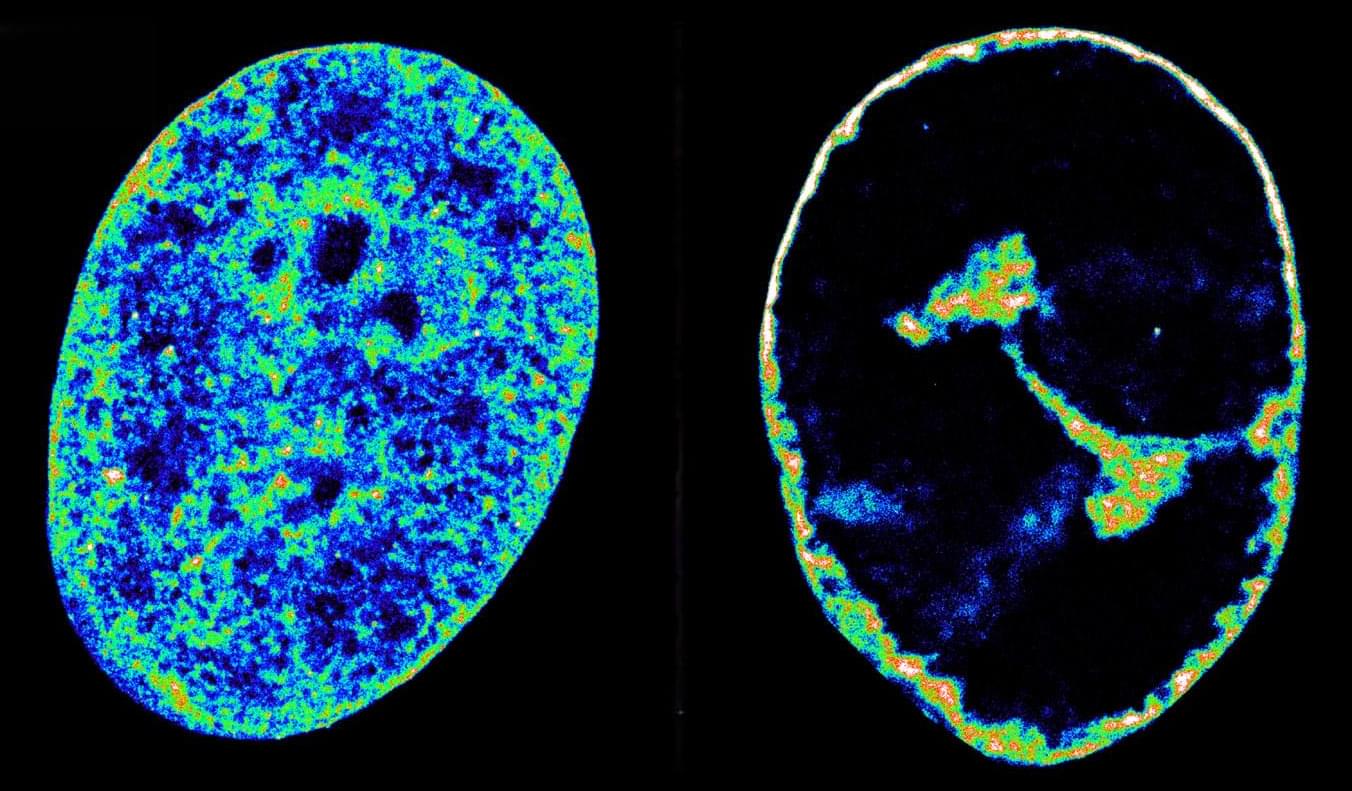

A study published in Cell Stem Cell reveals that some mutations in blood stem cells might help protect against late-onset Alzheimer’s disease.
A team led by researchers at Baylor College of Medicine discovered that both a mouse model and people carrying blood stem cells with mutations in the gene TET2, but not in the gene DNMT3A, had a lower risk of developing Alzheimer’s disease. Their study proposes a mechanism that can protect against the disease and opens new avenues for potential strategies to control the emergence and progression of this devastating condition.
“Our lab has long been studying blood stem cells, also called hematopoietic stem cells,” said lead author Dr. Katherine King, professor of pediatrics— infectious diseases and a member of the Center for Cell and Gene Therapy and the Dan L Duncan Comprehensive Cancer Center at Baylor. She is also part of Texas Children’s Hospital.
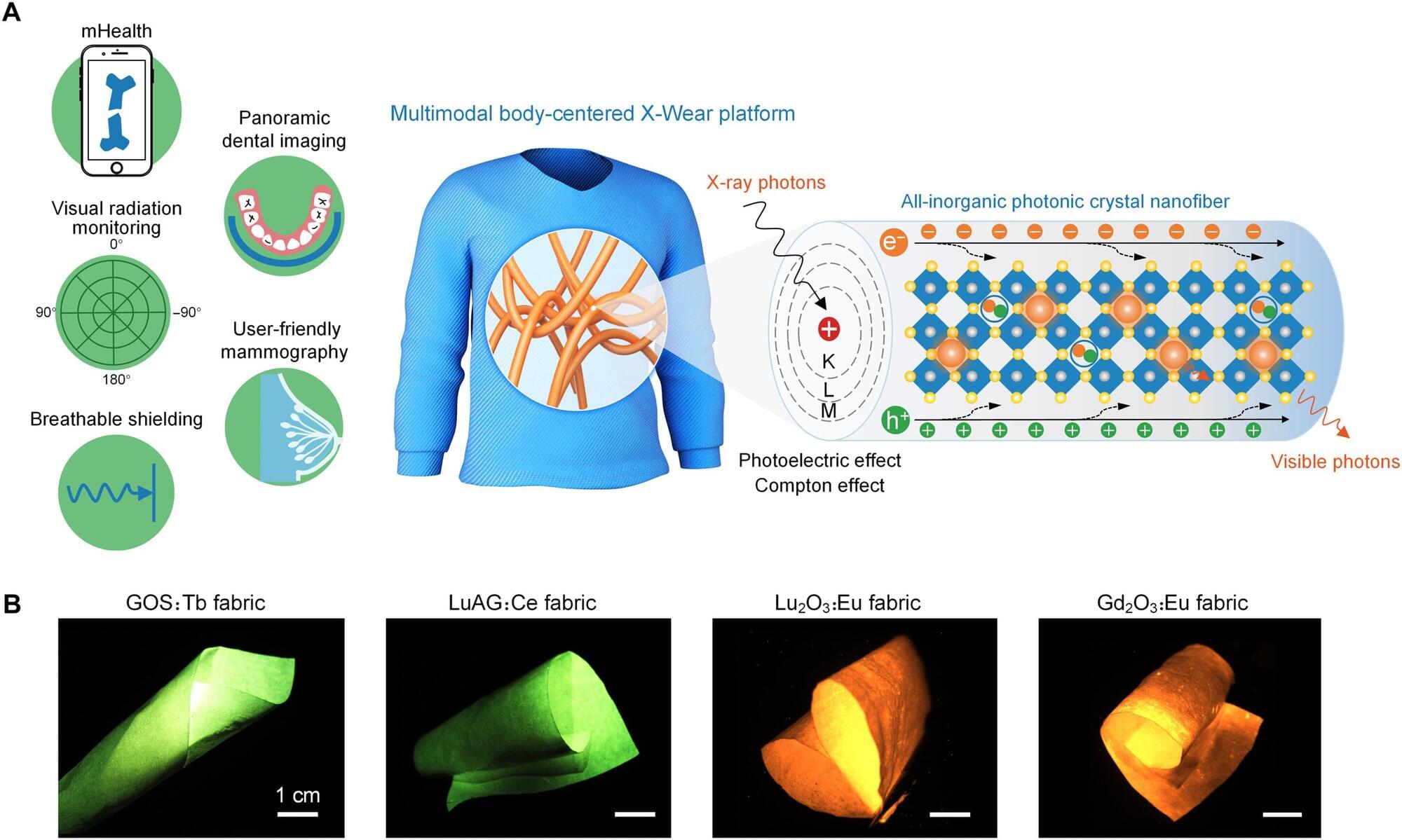
Since their discovery by Wilhelm Roentgen in 1895, X-rays have become a staple of modern medical care, from imaging teeth and broken bones to screening for the early signs of breast cancer.
The most common type of X-ray detector used in medical imaging today utilizes materials known as scintillators, which are made of inorganic and rigid compounds. This inherent lack of flexibility limits their applications and often requires patients to contort their bodies to accommodate unyielding medical equipment.
This rigidity has created a demand among researchers and the medical community for scintillating materials that are robust, efficient, and flexible. Past attempts to meet this demand, however, have had to sacrifice durability and efficiency for flexibility. An innovative fabric made of flexible inorganic fibers shows remarkable promise and may meet all three requirements.
{kind=link}