This video explores aliens, mind uploading to other species, genetic engineering, and future robots.
SOURCES:
• https://en.wikipedia.org/wiki/Eagle_eye#:~… https://www.scientificamerican.com/ar… • https://en.wikipedia.org/wiki/Human_c… ___ 💡 Future Business Tech explores the future of technology and the world. Examples of topics I cover include: • Artificial Intelligence & Robotics • Virtual and Augmented Reality • Brain-Computer Interfaces • Transhumanism • Genetic Engineering SUBSCRIBE: https://bit.ly/3geLDGO ___ This video explores the future of ChatGPT and 10 ways it could change society. Other related terms: aliens, alien species, advanced civilization, genetic engineering, robot, mind upload, mind uploading, brain computer interface, artificial intelligence, ai, future business tech, future technology, future technologies, etc. ℹ️ Some links are affiliate links. They cost you nothing extra but help support the channel so I can create more videos like this. #alien #aliens #avatar #avatar2 #geneticengineering #braincomputerinterface.
• https://vcahospitals.com/know-your-pe…
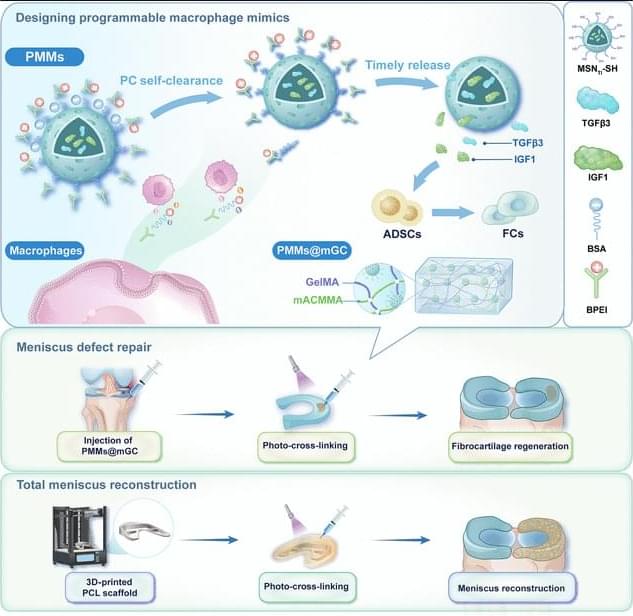
• https://www.scientificamerican.com/ar…
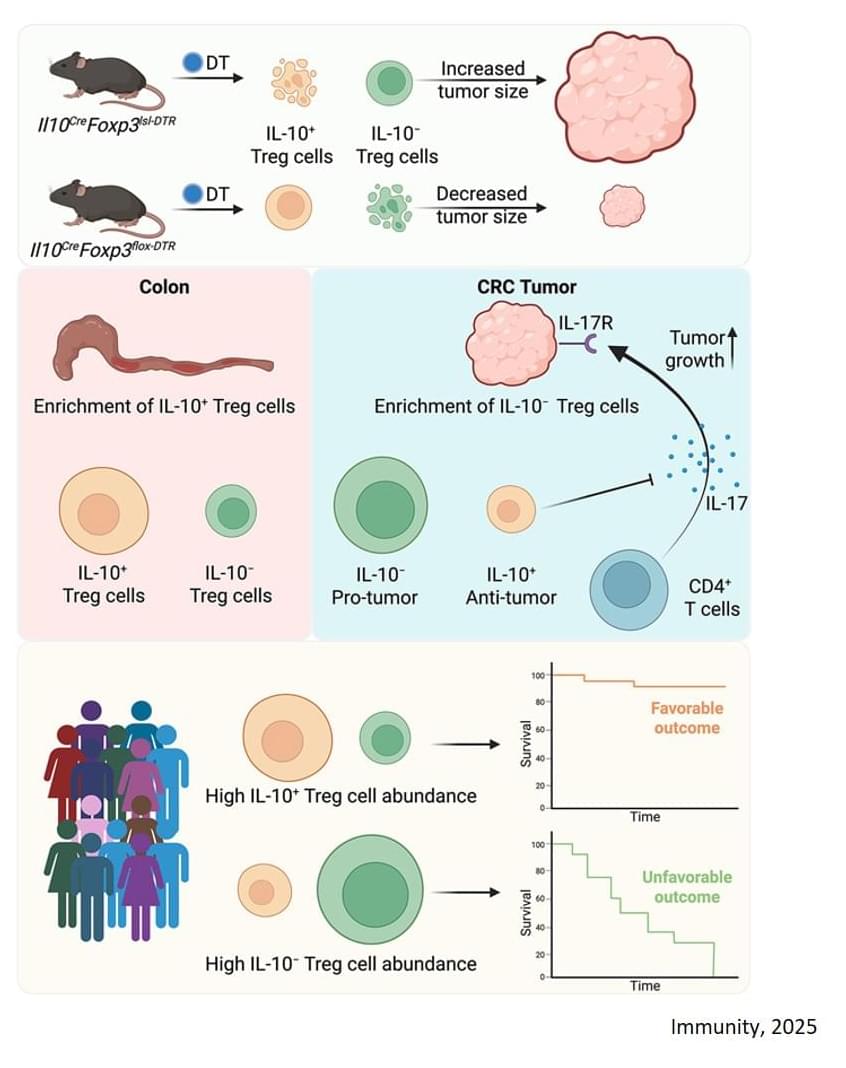
• https://en.wikipedia.org/wiki/Human_c…
💡 Future Business Tech explores the future of technology and the world.
Examples of topics I cover include:
• Artificial Intelligence & Robotics.
• Virtual and Augmented Reality.
• Brain-Computer Interfaces.
• Transhumanism.
• Genetic Engineering.
SUBSCRIBE: https://bit.ly/3geLDGO
This video explores the future of ChatGPT and 10 ways it could change society. Other related terms: aliens, alien species, advanced civilization, genetic engineering, robot, mind upload, mind uploading, brain computer interface, artificial intelligence, ai, future business tech, future technology, future technologies, etc.




{kind=link}