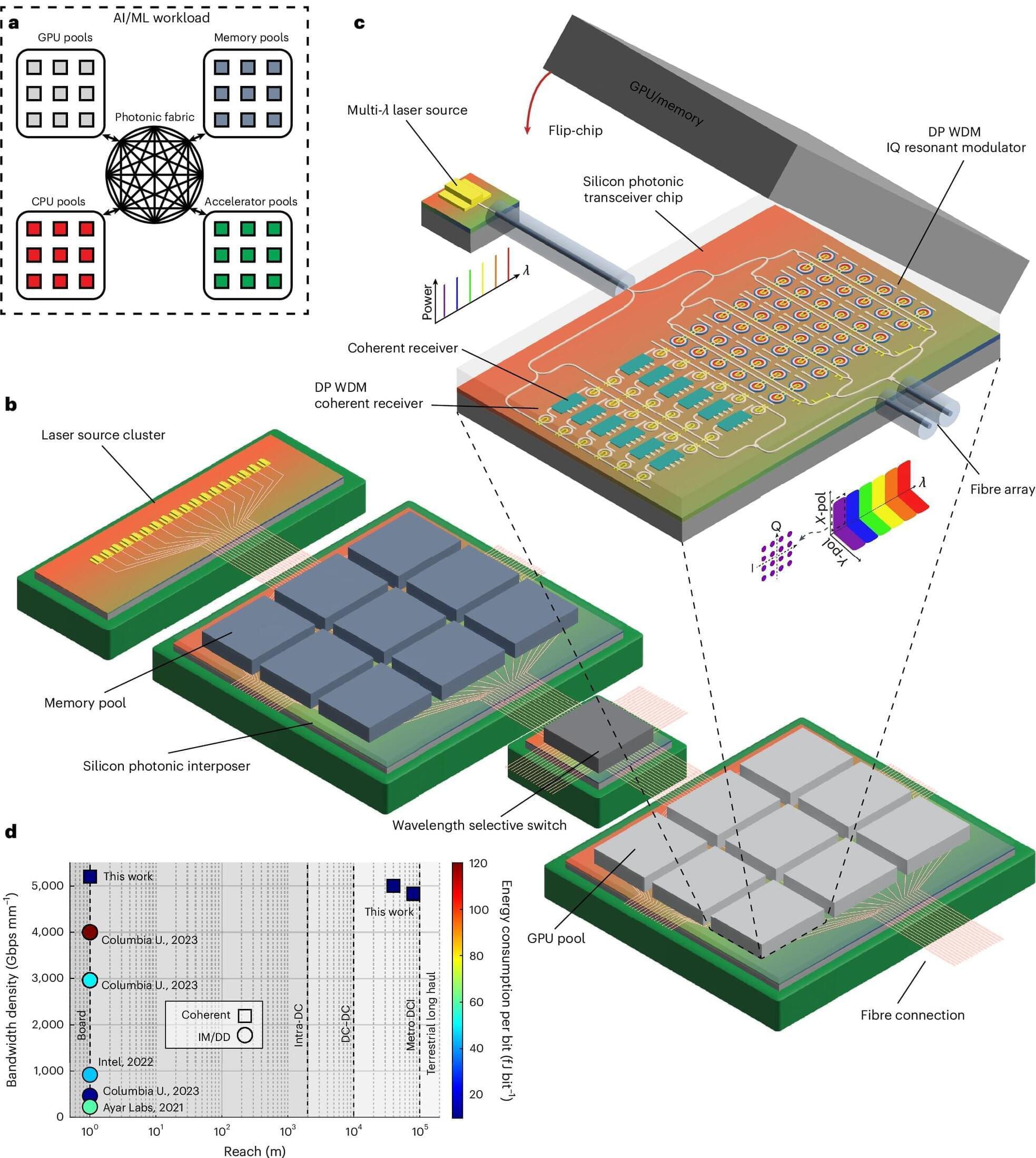
Artificial intelligence systems like ChatGPT are notorious for being power-hungry. To tackle this challenge, a team from the Center for Optics, Photonics and Lasers (COPL) has come up with an optical chip that can transfer massive amounts of data at ultra-high speed. As thin as a strand of hair, this technology offers unrivaled energy efficiency.
Published in Nature Photonics, the innovation harnesses the power of light to transmit information. Unlike traditional systems that rely solely on light intensity, this chip also uses the phase of light, in other words, its shift.
By adding a new dimension to the signal, the system reaches unprecedented performance levels, all while maintaining a tiny size. “We’re jumping from 56 gigabits per second to 1,000 gigabits per second,” says Ph.D. student Alireza Geravand, the first author of the study.




{kind=link}